

Vagaries

It takes two people to make you, and one people to die. That's how the world is going to end.
—Faulkner, As I Lay Dying
Only the blunt facts now, we’ll find the sharp ones later. Some things must be understood just to get them out of the way.
Here is what happened: once tree-bound creatures began to forage in the savanna, which put severe pressures on them, which led many to die, which led some few to conquer. First there was Australopithecus afarensis and then there were A. africanus and A. aethiopicus and then there were A. garhi and A. sediba and Paranthropus robustus and P. boisei and Homo habilis and H. rudolfensis then there were H. erectus and H. ergaster and then there were H. heidelbergensis and then there were H. naledi and H. neanderthalensis and H. florensis and H. denisovan and H. sapiens and then there is H. sapiens.
A Warning and an Overview
The following is a record of human evolution as it is understood now. I should say as I understand it now, not being a practitioner of any field involved or expert in anything vaguely related. In all cases, the mistakes and omissions are entirely my amateurish own.
This is not my normal style, later essays will be detailed and jump from time to time, I have my reasons for writing a schematic piece first. When I returned to write this after a few years off blogging it took me quite a long time to feel comfortable in any understanding of the hominin timeline. This was my own fault, and I would like to correct my errors before writing about the paleolithic for the next [number] of pieces.
I started reading about the paleolithic in a very piecemeal way. This was not a good plan. Taxonomy is incredibly contentious in paleoanthropology, tool classification equally so, phylogeny even more so. Since the same events and dates and specimen will be referred to and interpreted in opposite ways without warning, reading in a scattered way meant I crashed into all of that without the context to understand it. I then tried to rectify this with textbooks and popular overviews, and ran into another issue: there have been so many discoveries in the past few years that books cannot be published or revised quickly enough. These meet sometimes: once or twice early on I’d read several articles/books on the exact same bones but (1) they were assigned to different taxa and (2) the dates were completely different due to new methods or discoveries, and my brain would register the exact same hominin as three different ones across tens of thousands of years. You are probably smarter than I am. Hopefully, this helps anyway.
This essay, then, is a map to reference and refer back to, one with dates and names that are imperfectly described but are at least contextualized. In other essays, I’ll be extracting what I can from relatively small details, which can get one lost without some geography. It matters that, e.g., H. naledi was only 300 kya (thousand years ago) or so rather than 1 mya (million years ago), but it’s not immediately clear why unless you know what else was happening then. The brain fizzles out at such great time differences. Hence, a reference guide that has become slightly too unwieldy for its own good. This in no way aims to be comprehensive. It is just human cartography.
A note on taxonomy: I am going to try to identify differing names/opinions on names, but I’m inevitably going to make choices and there will be much more competent authorities that disagree with me for better reasons than I offer. At least half of those reasons have to do with teeth, which preserve better and make up more of the fossil record than practically any other bone. I will tell you that my bias is towards lumping, but that I am not perfectly consistent (sorry). In any event, here is a chart that shows the extremes between lumping and splitting those two, taken from Principles of Human Evolution:
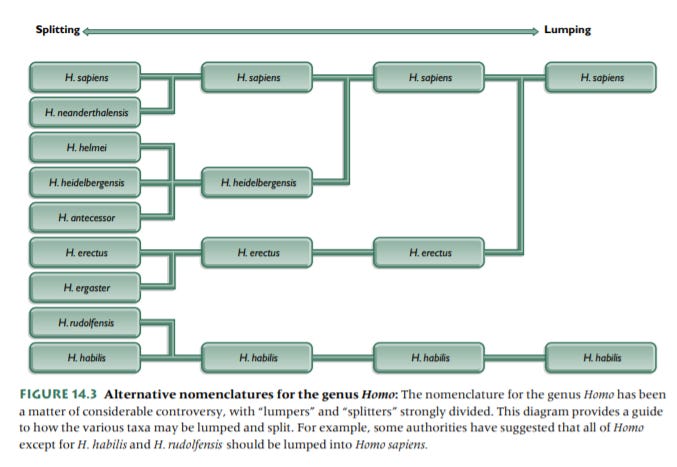
Ok.
This has a few themes. They will be revisited throughout, but it is worth noting up front.
Everything is much older than you think it is. It is, in fact, increasingly so. There are practically no new discoveries that have pushed dates forward. This goes for speciation, settlement, and technology. By the time I publish this, I fully expect some of the dates to be off by several hundred thousand years. To get the updates on all the ways this will proven wrong by someone who actually really knows what they’re talking about, Razib Khan’s genetics blog is excellent and The Insight is one of the only podcasts I actively listen to.
For a long time we seemed to really, really want there to be a single, simple line that led to Homo sapiens with minimal digression. But nature digresses. Human evolution is a story of contemporaneous species adapting and fighting for space, not our emergence at the tip of one great spear. We can easily forget this, as only Homo sapiens remains anymore. But Homo sapiens has been the sole human species since Neanderthals died out 39 kya (alternately, maybe 20 kya for Denisovans). That’s less than 1% of the standard “human evolution” timeline: 39000/4000000. To put that into perspective: the universe is 13.799 billion years old and wikipedia tells me that sturgeons first developed 245-208 million years ago. Taking the lower estimate, sturgeons have existed in the universe for a longer relative time (~1.5%) than humans have been a single species.
Along with that, there was (and is) a very common desire to tie human development to technology and brain-size and, by extension, brain-size to technology. But technology is just as chaotically classified as biology, and while there is progression in both, it is not as simple as “more-modern-looking-thing is so much better as to wipe out everything else instantly.” More to the point, human brain size is obviously important, but technology and brain size don’t track perfectly, and other things that really might matter - e.g. social learning - don’t leave precise fossils.
Interbreeding is among the most significant finds for interpreting modern humans’ recent past. Homo sapiens interbred with multiple other species, and there were almost certainly multiple hybridization events among those. This is very likely the case for earlier hominin, as well, making the already convoluted question of lines of ancestry even more so.
With this all out of the way, the story I am going to tell may often appear linear, which is inevitable. It is a very basic timeline, and I am lumping strongly at points. There is a strong probability that within a decade I will look back at this as a quaint attempt that used a limited understanding of antiquated findings. May we all hope so - it sounds like a good future.
Here is what happened in the deep past:
Pliocene Brave and the Unprepossessing Animal
In the Miocene (23.03-5.33 mya), apes began to fall from tree branch to tree base, and some of them began to move to the savanna. Around 8-10 million years ago (mya), gorillas split off from our line. Some 6 mya, the homini tribe splits into the subtribes Pan (chimpanzees and bonobos) and Australophithecine (our line). At the very end of the Miocene, these found themselves spreading out onto the plains and savannas of Africa. It is a slow process escaping the forest and it always has been.
Starting in the Pliocene (5.33-2.58 mya), the climate began to dry and cool and the Australopithecine start down a path that leads to me typing this and to you reading this. Ardipithecus ramidus, around 4.4 mya, is often cited as the real start to the whole show (the name ramidus - from ramid for “root” - is perhaps a give away). Ramidus still had adaptations for climbing trees, but ramidus walked - at least sometimes - bipedally, and ramidus was in the right place at the right time to point to Australopithecus. Their bones fossilize in the Afar triangle of East Africa, down where we began to find ourselves.
Around ~4-4.2 mya, Australopithecus anamensis begins the Southern Ape line. Though still primitive, it appears to have been habitually bipedal. Its jaw was still very ape-like (prognathic is the term), but its teeth are more human-like. Very quickly (perhaps 4.2 mya), we arrive at the most well-known of all australopiths - A. afarensis, known for the skeleton we call “Lucy.” Lucy comes, like ramidus and anamensis from the Afar region of Ethiopia. Specifically, she was discovered in the Awash valley, which is a fantastic name for a valley and its river. Others - e.g. the Laetoli collection - have been found in modern day Tanzania. Coupled with earlier bones of Anamensis discovered in Kenya, there was clearly a relatively wide a range for australopiths across East Africa.
With apologies to anamensis, Lucy serves as a pretty good introduction to the australopiths in general, and I will spend more time on her than the others. It is important, I say to myself as well, to recall that these were not just pre-humans. The australopiths were good at what they did, and what they did was spread across the land:
Overall, then, A. afarensis anatomy – and presumably behavior – is somewhat intermediate between that of an ape and a human, a pattern that does not exist today. As a result, many researchers tend to see the adaptation as “unstable” – that is, as being in the process of striving toward the “perfection” of the human model. In fact, this unique anatomical and behavioral repertoire should be considered a stable evolutionary package. After all, the species persisted for at least 1 million years, from 3.9 million to 2.9 million years ago, and possibly even longer. (Principles of Human Evolution)
But if that is the scientific truth, it is also important to recall why we may be so inclined to discount the species as stable. Drop one of us in the jungle and we’re helplessly outclassed, and Lucy was even smaller than we are. This was her world:
Without speed, claws, significant teeth, night vision, or protective coloration, these primates of 3.8 million years ago and earlier flourished. Walking across the same surface as the Australopithecus afarensis at Laetoli were tigers, rhinoceroses, elephants, and other animals that competed with the australopiths on these African plains, and one wonders how such an unprepossessing animal could survive in this environment. Perhaps they already possessed embryonic forms of the critical elements of cultural behavior: intelligence, protective human family relationships, and tool-use capabilities that were slightly better than that of the chimpanzees (who use sticks and unmodified stones). (Patterns in Prehistory)
The social aspect seems unquestionable, though it also applies to other terrestrial species, that “…face greater risk from predators than arboreal animals, [and so] live in larger groups and commonly include more males in the group; in addition, the males in such species frequently are equipped with large, dangerous canine teeth. (PHE)” But Lucy’s position is actually more interesting. Great Apes are substantially larger than our tree-bound relatives, with a common explanation being “fights.” Lucy is pretty gracile - about 3 1/2 feet tall, her males around 4 1/4 to 5 feet. In other words: there may have been other ways she survived.
A. afarensis’s cranial capacity was only slightly larger than that of a chimpanzee: 380-450cc compared to the chimpanzee’s 300-400cc. Some might think this implies an inability to create tools. I am less convinced. There is some evidence, though indirect, of rudimentary tool use we can associate with afarensis, and there are direct tools from the same time period, though they aren’t associated with afarensis. These tools -called “Lomekwian” - are from some 3.3 mya, and will be discussed in a separate piece. Still: they aren’t quite old enough to match the timeline, and the most surprising thing about them is almost certainly how quickly they seem to have come and gone.
Without tools, trees offer some safety lacking on the ground. Some theories posit that Afarensis may not have been exclusively terrestrial - her legs were quite short, which indicates small stride and tends to point towards greater arboreality. She also was not perfectly adapted for walking upright, and may have done so rather clumsily. Frankly, this question is part of a humongous argument I am not at liberty to resolve for you.
Afarensis’s teeth were certainly not built for fighting. It looks as though the teeth were adapted to long intense chewing of hard foods, but their microwear is most similar to the Gorillas that consume soft fruits and leaves, neither of which is known for being hardtack. Researchers allow for the fact that such food might have been fall-back, which presumably would have been a strong selector inasmuch as those famines would have lead to mass deaths. But then one would expect to run into a skeleton with some wear that indicates that - we have, after all, several hundred afarensis specimen. We also have them across time:
The consistency of dental microwear in A. afarensis postcanine teeth across time (Grine et al., 2006b) may mean either that these hominins were able to track their preferred dietary resources in the face of relatively subtle changes in the environment or that environmentally induced shifts in diet did not involve changes in mechanical properties of the foods typically consumed. (Lucy Redux)
This last discovery points to range. She either moved quickly and without great difficulty, or had some way of processing/handling the foodstuffs that allowed her to range and eat without cracking to pieces. This makes sense, because Lucy’s kind had already expanded well beyond East Africa.
A. africanus is contemporaneous with Lucy, emerging some ~3.6 mya in modern South Africa, best known for the famous “Taung Child” skeleton. They are different from Lucy (notably more arboreal), but are similar enough that we can generalize in the following way: for a moment, consider the sheer distance between Ethiopia and South Africa. Now consider what it would mean to traverse it. To help you picture it, forget about just hyenas and tigers. Those are terrestrial threats, and may be avoided in the way we avoid terrestrial enemies; the Taung Child died because their eyes were torn from their skull by a large bird. In addition, consider what the very act of movement itself might mean:
The tracks of two Australopithecus afarensis at Laetoli indicate that they may not have had quite the same smooth gait as modern humans do, as they seem to have a somewhat rolling walk in which the hips swiveled side to side more than they do when we walk. (Patterns in Prehistory, 102)
This should remind you of the way chimpanzees somewhat totter when they walk upright. Not exactly like that - she could walk better than a chimpanzee - but not quite as well as we can. But it was still - quite clearly - the preferred stance, inasmuch as she clearly was an upright ape rather than one that crawls. Were that not the case, we wouldn’t have tracks showing it.
To fall into tooth and sway and speciation is, at times, to forget what we aim to describe: the way of a small animal in the woods on the grass. Lucy walked upright as we walk upright, the first and last animal to stand tall. There were strong seasons in the Pliocene and the rains got heavy. She stayed by rivers for water, like all animals stay by rivers for water, and when there were enemies she hid in the trees or she died. There were many enemies. She could probably think more than a chimpanzee and there were hyenas then and one knows that once, at least, one of her kind was caught surrounded by their teeth without a tree to climb. She ate fruit.
In time Lucy’s kind was successful enough that they all died.
Pleistocene Teeth
Towards the very end of the Pliocene, one other important species emerges in East Africa: A. aethiopicus, first found around Lake Turkana, Kenya. They are also referred to as Paranthropus aethiopicus - the paranthrops are a contentious genus typically defined by robustness. The aethiopicus relationship with afarensis is uncertain, making the Paranthropus relationship with other genera uncertain. I don’t want to overstate the differences: they share many archaic traits, and some authors just reject Paranthropus entirely, but there are big differences relating to teeth. Aethiopicus is typically described as having a powerful jaw with very robust teeth (comparatively) - but here, as with afarensis, it is unclear what to make of microwear patterns that suggest relatively soft foods. Whether Paranthropus should be understood as a distinct genus is not up to me, but if they are then aethiopicus became the first of their kind around 2.7-2.6 mya, just before the Pleistocene, when the human saga gets bristly.
The Pleistocene begins 2.58 mya, and our major evolution may as well be synonymous with it. This is true biologically as well as technologically. At the very start of the Pleistocene, the first true archaeological industry emerges: the primitive “Oldowan.” But primitive does not mean “pick up a rock”, it means consciously craft artifacts through lithic reduction. You take a suitable rock in one hand (hammerstone), strike a larger rock (core) in the right spot, and the breaking results in sharp edges (here is a good video showing how to make Oldowan tools). It’s hard to explain quite what a leap this is. Chimpanzees use unmodified rocks as tools - they would take the hammerstone and be done. We take the hammerstone to another stone and then use the third stone. Human beings live in the future at all times, and our tools are the earliest evidence of that temporal dislocation.
The biological part is harder, if not simply because there is so much change. A panoply of hominins appear now, the most important of which is A. garhi, a small-brained australopith noted for being the first habitual tool-user and the creator of the Oldowan style. Straight from aethiopicus, two more important members of Paranthropus emerge. These, P. boisei and P. robustus, share the particularly large teeth of aethiopicus, the thick enamel of which appears to respond to some dietary change. This is in contrast with our line, whose teeth are notably small and feeble for such great apes. But the dietary change still occurred, and it’s a very telling one: “ evidence from microwear patterns on the surface of teeth shows that hominin diets remained predominantly vegetarian until approximately 2.5 million years ago, when cut marks are found on animal bones possibly associated with Australopithecus garhi.” (Principles of Human Evolution, 247)
This does not mean that everyone switched to eating meat. It does mean that the diet of hominins changes very suddenly, perhaps influenced by environmental factors, and meat-eating was one of the responses. There are two notable adaptations: one biological (robust teeth), the other behavioral (habitual tool use). Since we did not develop the teeth, it is hypothesized that our response (as in, the Homo line’s) came from the stone-tools camp. Presumably, this is aided by the knowledge that we still use tools.
But this does not explicitly tell us who our australopithic ancestors were. All hominin remaining in Eastern Africa seem to have gone down the robust-tooth line, excepting garhi, and garhi is almost certainly not our direct ancestor (for reasons that will take too long to get into). Perhaps more to the point: we keep finding homo-like bones that are older and older, as in “~500,000 years older” and thus couldn’t have come directly down from garhi.
I will relay two theories. One account, trying to account for our tooth discrepancy, argues that we are actually from a group of Africanus that migrated back into Ethiopia. The africanus that remained south, then, branched off into the relatively-recently-discovered Australopithecus sediba, which would make sediba a kind of sister species to our own line. However, recent discoveries of intermediate africanus seem to point to substantially more arboreality than even afarensis, as though we left the trees and then went back and then left again and all the while evolved traits to account for this in rapid succession. Seems unlikely. Hence, another account says that we likely emerged from something-else-but-still-actually-surprisingly-related-to-robustus: “…afarensis gave rise to an as yet unknown species that was aethiopicus-like in some ways (in traits not related to heavy chewing); this species was the common ancestor of aethiopicus on one hand, and gave rise to A. africanus, early Homo, and the later robust australopithecines on the other.” (303) You be the judge. Alternately: give me another theory.
Either way: the Homo line emerges around 2.3 mya (or this was the standard date, up until the very-very recent discoveries that pushed it as far as 2.8 mya - let me just finish this). The first specimen of our line, H. habilis, is also a habitual tool user, in the same mode as garhi. The relationship of H. habilis to the many hominins above is unclear, as is its relationship to the contemporary H. rudolfensis. Rudolfensis is much larger than habilis, but this may be a bias from the skeletons found (I’ve seen suggestions that we’re actually looking at incredibly pronounced sexual dimorphism - this seems unlikely and uncharacteristic for Homo, but who knows). I will be honest here: habilis is the most boring archaic human to me, despite the fact that it is the first of Homo. Perhaps this is because, frankly, the existence of Homo habilis appears to be a grudging admission.
Habilis was originally classified through behavior rather than anatomy - tool-use - and named for this: the “crafty man”. But the behavior is no longer unique. We began to find older and older assemblages of Oldowan tools, and now A. garhi is the presumed first tool user (garhi means “surprise”). As a result, the definition of the Homo line has more to do with teeth and cranial capacity than it does with behavioral leaps forward (habilis has a brain about halfway between Australopiths and Homo erectus).
Hence, what strikes me as genuinely interesting are not the specimens but the interaction. Garhi constructed tools in the Oldowan style, and habilis adopted Oldowan tools later. But if garhi is not the direct ancestor of habilis, that means that it didn’t pass Oldowan down directly while it adapted itself out of existence. And if habilis didn’t independently create the same exact industry, then habilis must have learned Oldowan crafting from garhi.
Give me this “if”: the old linear progression of australopithecus implied a direct familial tradition of technology. This more chaotic version - garhi and habilis living alongside one another - points to overlapping relationships between separate human species. We - here speaking as Homo - are not so defined by our individual inventiveness as we would like. The more ubiquitous and central capacity of the human species is, and always has been, a heightened ability for the individual to learn.
Losing the “if”: there are arguments that Oldowan styles are simple enough to be independently developed over and over, and thus we should not intuit complex social learning from stone industries. Remember that I am not an authority on any of this, but I will tell you that I find this argument unpersuasive. Two reasons: 1) we find evidence for the importance of social-learning in chimpanzee tool use for substantially simpler technology than Oldowan (also: behaviors in general); 2) a consistent theme of technologies is their loss when proficient members of the group die out or disappear and no one can properly teach the youth, true for both humans and apes. See, for example, the Lomekwi tools.
Now things begin to move very quickly.
Up Man
We spread from Africa early.
For a long time it was thought that Homo erectus came directly from habilis, but this is now disputed. Part of the issue lies with the habilis/rudolfensis distinction. Another issue comes from the fact that erectus is not just later-habilis, but overlapped with habilis for perhaps ~300,000 years. And then there’s something of a geographical mystery. The hominins thus far described are centered in either Eastern Africa or Southern Africa. Erectus, however, has been found at practically simultaneous dates in Southern Africa (2 mya) and China (2.1 mya). If these are, in fact, both erectus, then it’s unclear where it first “originated.” Quite rapidly, we see erectus in Dmanisi, Georgia, some 1.85-1.7 mya, and in Java around 1.5 mya. There is no solution here for you - I genuinely do not know where erectus is from, but if it’s not from Africa then that means an earlier member of Homo made it across the Sinai.
The Acheulian tool industry, and its characteristic hand axe, is attributed to Erectus. That hand axe is the single longest-used piece of technology in human - and, barring new discoveries, universal - history, in continuous use for ~1.7 million years. In general, the Acheulean industry began what you and I would really recognize as stone tools: care taken to produce crafted, symmetrical tools by working both sides equally. This may have involved sharpening with materials since decayed: bone, wood, hide (this last may have been a later innovation). It’s not unlikely that many other tools in use are unknown to us for the same reason: stone tools have been found almost everywhere we’ve found H. erectus, but South/Southeast Asia have practically none. One hypothesis (tested, in a sense), is that H. erectus in S/SE Asia primarily used bamboo tools which have long since disappeared.
Unsurprisingly, tool use is reflected biologically: a relatively recently uncovered bone shows that H. erectus had similar adaptations, and - by hypothesis - was well adapted to tool use and already had modern-ish capacity for tool manipulation. Back to teeth:
A major shift occurs, however, with Homo erectus, whose enamel is heavily pitted and scratched. Such a pattern resembles a cross between a hyena (a bone-crunching carnivore) and a pig (a rooting omnivore). Although it is not yet possible to interpret precisely the implications for the Homo erectus diet, it is significant that tooth wear patterns indicate some sort of abrupt change in hominin activities at this point in history – perhaps significant brain expansion, reduction in body-size dimorphism, systematic tool making, use of fire, or migration out of Africa. (Principles of Human Evolution, 191)
Since I am not a scientist and I feel comfortable connecting some things, it seems like not-a-coincidence that we adopt “bone-crunching” teeth as our diet becomes much more meat based. Feel free to yell at me in the comment section. I will also observe that must be understood alongside the suggestion that our evolutionary course from the australopiths can be charted from more gracile teeth (the from-africanus one above), inasmuch as tool-making and new teeth and meat-eating seem to go together. Perhaps this was enabled - or driven by, or itself drove - the much, much larger brain of H. erectus: compare the ~600 cubic centimeters from H. habilis to the ~950 cc of H. erectus to modern humans’ ~1250cc. I have varying degrees of skepticism when reading into theories that put brains in the evolutionary cockpit, more on that later. For now, note that over the course of its existence, erectus’s brain seems to grow and its skeleton starts looking more and more modern.
Most interesting are what we can extrapolate sociologically. Sexual dimorphism is reduced as compared to the australopiths, indicating what is likely a more cooperative mating strategy among groups (likely a form of monogamy or near-monogamy). Erectus also begins to show a longer childhood - in us, that indicates cultural development and deep social learning. It also indicates cooperative raising. H. erectus shifted his weight like a modern human, and note that we’re adapted for long-distance running and travel. Those distances are aided by human social dynamics. A father or mother or both may wander out to hunt and forage provided that others care for their young. Humans are known to forage for multiple days, which is possible because our groups are not merely familial. Unrelated members of the same tribe watch over the collective young when away. Further, H. erectus was the first of us to domesticate our best and oldest ally: fire. Like all domestication events, it has the double effect of domesticating the domesticator. Anyone who’s been camping understands the need for group solidarity in maintaining a fire. A tender stays with the living flame and he must know that a hunter brings his food; the hunter has to know a hearth will be there when he returns.
In short: erectus began to behave like us. The strategy worked.
It worked spatially, which you can tell from just how many distinct “species” have been separated from and then woven back into erectus by excited scholars: H. ergaster, the early African variant; H. pekinses, “Peking man” in China; H. georgicus, the erectus found in Dmanisi… H. antecessor, the furthest removed, first appears in Europe around 1 mya (it is unclear whether we should call this erectus, but know that it sometimes is and appears contemporaneously).
And it worked temporally. Erectus overlapped with not just our ancestors, but with modern homo sapiens. Likely the longest-living hominin, erectus enters halfway through an essay written almost entirely in “million years ago (mya)”. The final erectus - once called Homo soloensis for the Solo river in Java - leaves about 100 thousand years ago (kya).
The Out-of-Africa hypothesis is now ubiquitous, and has been since the 90s. It is correct, inasmuch as modern humans only appear to have split from one another around 120 kya at the very outer edges, and most modern humans split around 60 kya when a few began to scramble across the Sinai. But there was a lively debate before between Out-of-Africa and the opposing theory of multiregionalism. According to the multiregionalist hypothesis, human beings evolved in multiple areas simultaneously, the result of (presumably) convergent evolution from an earlier hominin. This strikes us as insane now, but it must be understood in the context of so many similar bones spread so far apart. H. erectus - or the various hominins that make H. erectus, should you split them - is practically everywhere. Speaking naively and bluntly, there is something much stranger about the idea that all of these strands just died out, that the creature spread so far and wide and brazen would falter, that a relative which hadn’t left home before the 100,000s would usurp its place.
I understand the multiregionalists. It is hard to admit that the world of H. soloensis is a just and coherent world.
The Last of Others
At the exact same time as whatever-was-we began to converge on what-we-are, some primitive man called Homo heidelbergensis set out to prove the multiregionalists correct in a very limited sense.
I do not know if that small group - we, H. sapiens - ultimately came from erectus. It is now considered to be unlikely that they’re our grandparents. A more robust hominin called H. ergaster was often tapped for the sapiens line, but then again, ergaster may also simply be another way of saying “erectus”. Either way: the relationship between erectus and other Homo is, so the refrain goes, uncertain. What is certain is that the later heidelbergensis happens to fill the gap between H. erectus and three interrelated species: H. sapiens, H. neanderthalensis, and H. denisova. In other words: he’s almost certainly your closest ancestor, or something like it, depending on how you split the species, on how close you are to your grandparents, on how the fossils fall. At the very least, it is the closest thing we know to our parent species.
H. heidelbergensis is not well known, which is a shame. He first appears in Europe some 800-700 kya, just when antecessor disappears, and perhaps 700 kya splits off into two separate groups. One, in Europe, gives us the name “heidelbergensis”; the other, found in Ethiopia, Zambia, and South Africa, is called H. rhodesiensis. These are largely considered to be the same species, but if you see rhodesiensis, now you know who it is. That split is particularly important for the following reason: it is likely that the Eurasian variant gives rise to neanderthals and denisovans. Genetically speaking, the Homo sapiens branch is removed from that of H. neanderthalensis / H. denisovan by around 770,000-550,000 years. In other words: if heidelbergensis is the forefather of those two branches, then however and whenever he emerged, one group quickly wound up in Europe while the other wound up in Africa.
Many of the best finds of heidelbergensis are in caves, which makes sense: H. heidelbergensis looks like what you imagine a caveman would look like. Thick-skulled and heavy set. Anatomy that grunts. An individual found in Boxgrove Lagoon, England, was 5’11” and 200lb of muscle, in other words a strong and modern man, and he used his jaw as a vice while he cut. He hunted big game, the elephants and rhinos and hippos and horses that once populated Europe, with unambiguously hafted tools (Clactonian industry). Edges are notched, and a wooden shaft has been preserved. Short-range ambush hunting, presumably with spears. This seems, I suppose, simple. But I would like you to pause for a moment and truly consider what it means for a human with a bit of wood to fight an elephant.
“Anatomy that grunts” but aesthetic judgments tell us little. Heidelbergensis cared for the old and the dead. He continued to whittle down into human features: entirely upright, hairless, craftier, adapted for handedness, larger brains, and low-levels of sexual dimorphism. It is not unlikely that they had ritual burials of some kind, though the evidence for that in caves is conflicted. A ritual-looking hand axe appears along with the bones of their dead; the extent to which we can understand this as a grave-offering is for a later piece. From damage to the teeth, we can tell that Heidelbergensis liked toothpicks.
Towards the end, the skulls begin to look more and more neanderthalic. Bones from the Sima de los Huesos in Spain, dated to around 430 kya, are alternately referred to as heidelbergensis or neanderthalensis depending on the author. Note the timing. From genetic evidence, we know that neanderthals and denisovans split from one another between 470 and 380 kya. Neanderthals mostly inhabited Europe and parts of Southwest Asia. Denisovans pushed further East, and the most recent suggestion is that they were most plentiful in Southeast Asia and Sahul from at least 200 kya (nevertheless, they were everywhere from Tibet to Siberia). I will be writing more about neanderthals/denisovans in detail, because they occupied and continue to occupy the parts of my brain that interest me in history at all. But here, it is perhaps best to observe just how similar they are to us and the ways in which they are different.
In Europe, neanderthals emerge around 300-250 kya, with the “classical neanderthal” taking shape some 150 years later. This shape is much like you recall: a large nose, heavy-set shoulders, much more robust teeth and jaw, shorter, a kind of boxiness to the body. For a long time, likely because of this, there was an assumption that neanderthals were just-about-human but defective-in-some-way. Still tethered to various “Great Leap Forward” hypotheses, a lot of theories assumed that neanderthals lacked the linguistic or social or cognitive complexity to go toe-to-toe with H. sapiens. One of the ways this comes up, still, is in the study of stone tools: classification of tools for some time merely assumed that the more complex ones came from H. sapiens, and were then used to date sapiens’s entrance into neanderthal territory. This is wrong.
We now know that neanderthals could speak like we can speak (albeit with a different tenor) - their faces are shaped correctly, they have the right FOX-P2 variant (if that matters at all), their hyoid bone is ready. Their brain was slightly larger than ours, on average. More to the point, neanderthals created obviously symbolic artifacts extremely early on. 176,000 years ago we find what I’m comfortable describing as a “shrine” of buried deep in a cave. Some 115-120 kya we find neanderthal jewelry in Spain. At 65 kya, we find the first painting anywhere, a neanderthal masterwork.
We have substantially more limited information on denisovans, simply due differences in the fossil record. But given their close relationship to neanderthals, it seems likely that they were comparably advanced. Indeed, it is unclear how much we can separate the three heidelbergensis lines - neanderthals also inhabited parts of Asia, and first generation neanderthal/denisovan hybrids have since been found. Recovering a 50/50 split seems astronomically unlikely if interbreeding was very rare. To foreshadow some: we also interbred with neanderthals and denisovans, hence the limited multiregionalist triumph.
But this is all too linear. Given this focus on the heidelbergensis-esque line, one almost forgets how many other archaic humans were around. In Southern Africa, an entirely different species lived: H. naledi, a much smaller and more ape-like hominin that may or may not have buried its dead in caves. Though no tools have been found, it likely had Acheulian artifacts. In Flores, modern Indonesia, a species called H. floresiensis lived, known colloquially as the “hobbits.” At some point in there, denisovans, or neanderthals, or h. sapiens, or all three of us, interbred with a group that David Reich calls “Super-Archaic Hominin,” known by its divergent genetics and nothing else. And, of course, erectus was still puttering about Java.
All those species we did not interbreed with - naledi, floresiensis, erectus - are notable, in a sense, for their small brains. But this is only notable if we assume that large brains are such an exceptional advantage that evolution practically wants and aims for them, will wipe out anything nearby with a slightly smaller skull. There is little reason to think this. Small-brained hominin held on for some time and used tools that weren’t so dissimilar to their contemporaries. Brains are not everything. Neanderthals have slightly larger brains than modern humans. Look what happened to them.
Modern Alone
The first modern human bones are in Jebel Irhoud, Morocco, 300 kya. H. rhodiensis - the African variant of heidelbergensis - disappears at around this same time. There are just-about-simultaneously collections found in the Rift Valley, Kenya, that point to incredibly “modern” behavior, including pigment production and obsidian distant enough to suggest trade networks. Once, I think, it could be maintained that some kind of “great leap forward” in human behavior took place some 60,000 years ago or so. I don’t particularly understand how one can keep that up now.
Because we were first found in Morocco, it is genuinely unclear where exactly H. sapiens developed. The vast majority of our fossil record is still centered in East Africa/North Africa, and it is not terrible to imagine that to be the case. Based on genetic evidence, it seems that we remained together until relatively recently. As noted above, the last common ancestor for modern humans - mitochondrial Eve - is dated to ~120 kya. At that point, a group ancestral to the modern Khoisan separated from the rest of the human line. Two attempts at leaving Africa, and modern Homo sapiens begin to spread.
The first is a failure: perhaps 100 kya, one group of sapiens left Africa and began settling the Levant. While it’s not clear exactly what happened, this was a failure. Modern humans soon disappear and Neanderthals begin to inhabit the same region. My sense is that we were beaten back, that there may not have been enough of us and were more of them. This is speculative. It is hard to tell.
The second push succeeds: a new group of humans tries the same thing perhaps 70-60 kya. This group likely cohabited the middle east, for at least some period, with neanderthals, but before long neanderthals begin to disappear from the middle-east. At around this same time period, we likely interbred with those neanderthals. That group is ancestral to every non-African human on earth, and it would account for why neanderthal DNA (up to 2%) is present in every human population outside of Africa. If it happened elsewhere, the timing is all wrong, because we swiftly spread across the globe.
Modern humans colonize Southeast Asia and then Australia almost immediately, between 65-50 kya. At this point, we begin making our way all throughout Asia, and somewhere in there modern groups across Asia wing up with denisovan DNA (perhaps .5-1%), as well as one or two positive denisovan adaptations for high-altitudes. Another group of modern humans remains in the Middle East, and a third arrives in Europe around 10,000 years later (~41 kya). Recent research has pushed human habitation of the Americas back substantially. It is now possible that we first arrived in the Americas 33 kya. This is followed by a wave no earlier than 25-20 kya which includes the ancestors of most modern Native Americans, and which expanded throughout both continents by 16 kya. By that point, then, H. sapiens lives on every continent but Antarctica, in practically every modern country, and has dominated the local environments.
It is unclear when H. naledi and H. floresiensis died off. The first is dated to 300-200kya, the latter to perhaps 100-50 kya. Some time after that, then. It is also unclear whether they interacted with modern men, although it would surprise me if naledi did not. By 38 kya, only a few thousand years after we enter Europe, neanderthals are extinct. The final denisovans lived in Sahul, the old continent that we now know as Australia-through-New-Guinea. These last denisovan must have made it to at least 40-38 kya to account for their genetic contribution; there was more interbreeding in Sahul, leading to notably higher levels of denisovan in modern Papuans (up to 5%). Some researchers argue for a final date of 20 kya. If so, they lasted long enough to see us conquer the globe. We will not know what happened to them. I suppose it depends on what you think of modern men. Let us guess, then, that by 30,000 years ago, H. sapiens are that most modern adverb: alone. The ice has begun to push us in, fire pushes the beasts out. 4 million years of branching evolution, overlapping and interbreeding, struggling to escape its first continents, then dying and falling as other lines do, has terminated.
Here is what happened: tree-bound creatures began to forage in the savanna, which put severe pressures on them, which led many to die, which led some few to conquer. First there was Australopithecus afarensis and then there were A. africanus and A. aethiopicus and then there were A. garhi and A. sediba and Paranthropus robustus and P. boisei and Homo habilis and H. rudolfensis then there were H. erectus and H. ergaster and then there were H. heidelbergensis and then there were H. naledi and H. neanderthalensis and H. florensis and H. denisovan and H. sapiens and then there is H. sapiens.
Man carves sculptures of lion men, dyes the walls of his den, paints the bodies of his dead and plants them in the dirt.
We make flutes of bone and we play them when it is too quiet.

So glad to see you writing and publishing again. I hope you are well.
Genuine question, how does this link with your samzdat posts? Just curious